
Non-classical Major Histocompatibility Complex (MHC) molecules are non-polymorphic proteins that are implicated in both the innate and adaptive immune response.1,2 While classical MHCs present antigens to CD8+/CD4+ cells for further immune processing, non-classical MHCs also mediate activation or inhibition in natural killer (NK) cells.3 MBL International has your non-classical MHC needs covered. We offer MHC tetramers against key targets- HLA-E and MR1 (human), and Qa-1b (mouse).
HLA-E
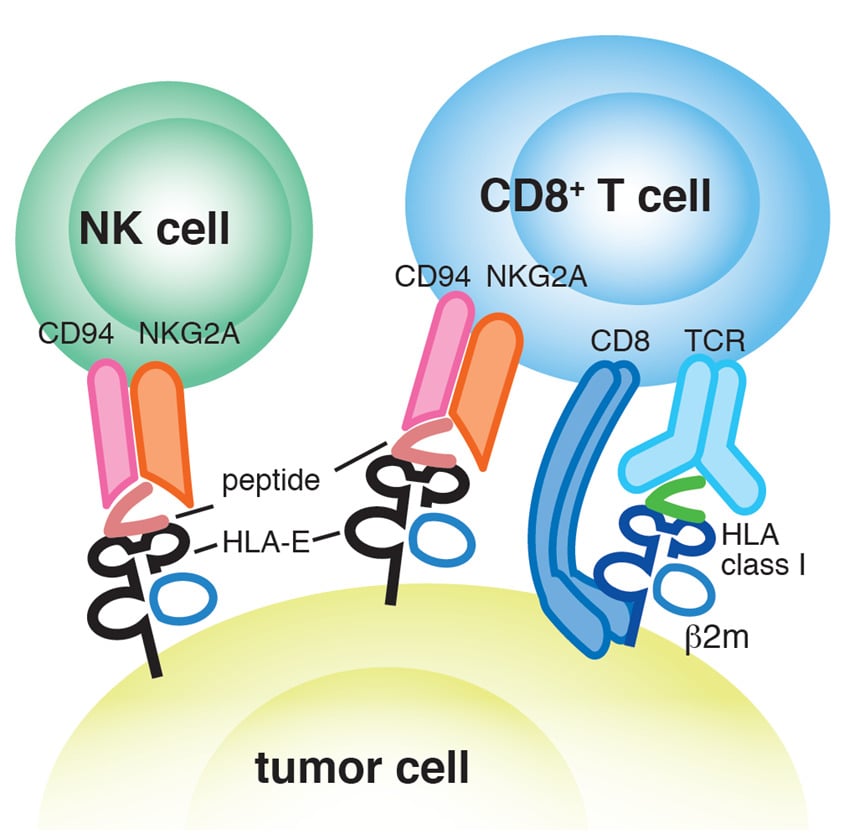
HLA-E is a non-classical MHC Class 1b molecule, expressed ubiquitously in humans. Two functional HLA-E alleles have been observed- HLA-E01:01 and HLA-E01:03. These molecules are recognized by the CD92/NKG2 receptor on NK cells and the T-cell receptor (TCR) on NKT cells. CD92/NKG2 receptor recognition allows for modulation of cytotoxicity and cytokine release by NK cells. Additionally, HLA-E molecules are involved in presentation of antigenic peptides to CD8+ T-cells through the αβ TCRs.4
MBL International offers HLA-E tetramers comprised of human class I HLA-E01:03 and epitope peptide derived from the HLA-A leader. The tetramer allows for detection of HLA-E01:03-restricted HLA-A leader3-11-specific NK cells and CD8+ T cells by flow cytometry.
Qa-1b
The Qa-1b molecule is the murine non-classical MHC class 1 homolog of HLA-E. It is characterized by limited polymorphism and is similar in function to the HLA-E molecule.5
MBL International offers biotinylated Qa-1b custom monomers and tetramers available tagged with PE, APC or BV421.
MR1
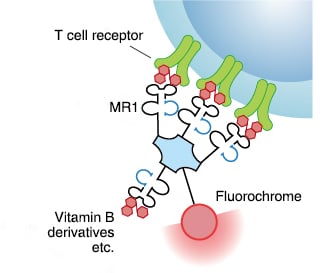
The MHC class I related protein (MR1) is a non-classical MHC molecule that presents ligands to the mucosal associated invariant T (MAIT) cells. These molecules are membrane proteins that are non-covalently bonded to β-2-microglobulin (β2m). These molecules are located in the endoplasmic reticulum. They migrate to the surface and present unusual ligands, such as vitamin metabolites from certain microbes to MAIT cells, thereby activating them.6
MBL International offers MR1 tetramers which are prepared by tetramerizing biotinylated human MR1/β2m complexes with streptavidin.
CD1d
CD1d is a membrane protein found in human and mouse species. The protein is non-covalently bonded to β2m and mediates the presentation of self and microbial lipids and glycolipid antigens to T-cells.7 The CD1d-restricted NKT cells are not only implicated in the activation of innate and adaptive immune responses, but they also express effector antimicrobial molecules.8
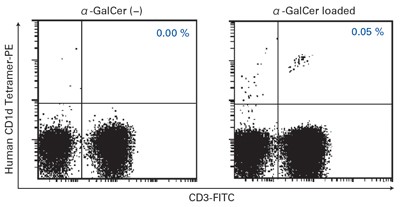
MBL International offers CD1d tetramers. These prepared by tetramerization of complexes of CD1d and β2m by PE- or APC- labeled streptavidin. Binding this reagent to α-GalCer enables highly sensitive detection of CD1d-restricted NKT cells and can be combined with antibodies to study NKT cell function through flow cytometry. α-GalCer loaded and α-GalCer unloaded tetramers are available.
For Research Use Only. Not for use in diagnostic procedures.
Learn more about the other types of MHC Molecules
Class I
MHC Tetramers- Human and Mouse Class I

Class II
MHC Class II Tetramers

References
- Blumberg RS American Journal of Physiology 274(2): G227-G231 (1998)
- Braud VM, et al. Current Opinion in Immunology 11(1): 100-108 (1999)
- Halenius A, et al. Cellular & Molecular Immunology 12: 139-153 (2015)
- Socié G, et al. Immune Biology of Allogeneic Hematopoietic Stem Cell Transplantation: 19-38 (2013)
- Oliviera CC, et al. Journal of Experimental Medicine 207(1): 207-221 (2010)
- Krovi HS, et al. Immunogenetics 68: 549-559 (2016)
- Dutronc Y, et al. Tissue Antigens 60(5): 337-353 (2002)
- Sköld M, et al. Infectious Immunology 71(10): 5447-5455 (2003)
- Miller JD, et al. J Immunol 171: 1369-1375 (2003)
- Braud VM, et al. Nature 391: 795-799 (1998)
- Lee N, et al. PNAS 95: 5199-5204 (1998)
- Tomasec P, et al. Science 287: 1031-1033 (2000)
- Heinzel AS, et al. J Exp Med 196: 1473-1481 (2002)
- Pietra G, et al. PNAS 100: 10896-10901 (2003)
- Salerno-Gonçalves R, et al. J Immunol 173: 5852-5862 (2004)
- Derré L, et al. J Immunol 177: 3100-3107 (2006)
- Monaco EL, et al. Neoplasia 13: 822-830 (2011)
- Weder P, et al. Results in Immunology 2: 88-96 (2012)