
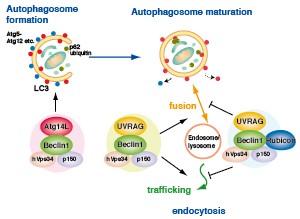
Atg14, Beclin1, Rubicon and UVRAG
Atg14L forms a class III PI3K complex with Beclin1, Vps34 and p150. The Atg14L complex has been shown to accumulate in the isolation membrane and to function as a positive regulatory factor for autophagosome formation in the early stage of autophagy. Moreover, it has also been demonstrated that localization of Atg14L occurs not only in the isolation membrane and autophagosome but also in the endoplasmic reticulum membrane, suggesting a possible important role of the endoplasmic reticulum in autophagosome formation.
The complex consisting of Beclin1, hVps34, p150, Rubicon and UVRAG has been reported to regulate the processes of autophagosome/lysosome fusion (Autophagosome maturation) and endocytosis. These finding imply that the two important cell functions, autophagy and endocytosis, may be regulated by Atg14L and Rubicon via Beclin1.
Antibodies
ATG16L
Atg16L forms an approximately 800 kDa complex with the Atg12-Atg5 conjugate. The Atg16L complex is localized in the isolation membrane (also called the phagophore) under starvation conditions, and facilitates the conjugation of LC3 to PE. Lack of autophagosome formation in Atg16L knockout mice clearly indicates that Atg16L is an essential factor for autophagy.
Atg16L shares high homology with yeast Atg16, but unlike yeast Atg16, carries a large WD repeat domain at the C-terminus. In the WD repeat domain, a sensitive single nucleotide polymorphism (SNP) has been detected for Crohn’s disease, one of the inflammatory bowel diseases of unknown cause. The association of Crohn’s disease with Atg16L is a topic of further study.
Anti-Atg16L antibodies:
Code No. M150-3 (Monoclonal) & PM040 (Polyclonal)
Atg9 and Atg-2Atg18 complex
Atg9 is not known where the initiation of autophagy takes place, but in yeast, autophagosome formation is suggested to start at the pre-autophagosomeal structure (PAS). Atg9 is thought to be involved in the early phage of PAS formation, since many Atg proteins are not localized in the PAS in the budding yeast lacking ATG9. Atg2-Atg18 complex binds to Atg9 on the surface of the PAS. Atg18 is known to interact with PI3P, however, the function of Atg2-Atg18 complex remains to be elucidated. In the yeast with mutated ATG18 and ATG2, Atg9 is highly accumulated at the PAS. Meanwhile, Atg9 is not localized at the surface of autophagosome. These finding suggested there must be mechanisms for Atg9 to detach form the PAS surface. Atg2-Atg18 complex may be involved in this process.
Anti-Atg2A antibody:
Code No. PD041 (Polyclonal)
Anti-Atg9A antibody:
Code No. PD042 (Polyclonal)
ULK1-Atg13-FIP200 complex
ULK1/Atg1 is a serine/threonine protein kinase essential for autophagy. In mammals, ULK1 usually forms a large complex with Atg13 and FIP200, which are highly phosphorylated by the serine/threonine protein kinase known as mammalian target of rapamycin (mTOR), leading to its inactivation. Under starvation conditions, mTOR dissociates from the ULK1-Atg13-FIP200 complex, resulting in reduced phosphorylation of ULK1 and Atg13. This triggers localization of the ULA1-Atg13-FIP200 complex in the isolation membrane and recovery of ULK1 kinase activity, thereby promoting autophagy.
Anti-Atg13 antibodies:
Code No. M183-3 (Monoclonal)
Learn more about other popular Autophagy targets and products
Autophagy
Autophagy is a cellular self-digestion process for the purpose of providing nutrients to allow for cell survival during stress conditions.

LC3/ATG8
LC3 is most actively studied and frequently used as a mammalian autophagy marker.

P62/SQSTM1
p62/SQSTM1 is thought to be a substrate that binds directly to LC3 causing its selective degradation by autophagy.

ATG12
Ubiquitin-like protein Atg12 has a C-terminal glycine, which protects it from processing.

NAD+ Biosynthesis
NAD+ Biosynthesis Assay Kits
